
Existen tres categorías de
elementos formes o elementos figurados:
- Los eritrocitos, hematíes, glóbulos rojos o células o corpúsculos rojos de la sangre.
- Los glóbulos blancos, leucocitos o células o corpúsculos blancos de la sangre.
- Las plaquetas o trombocitos.
Eritrocitos:
Los
eritrocitos son discos bicóncavos no nucleados (no poseen núcleo), con un
diámetro medio de 8.5 micrómetros con un espesor en los bordes de 12
micrómetros y en el centro de un micrómetro. Esta forma es la más ventajosa
porque representa la superficie máxima en relación a su tamaño para la difusión
de gases.
 Los eritrocitos maduros carecen de núcleo pero tienen metabolismo,
consumen O2, ATP y glucosa y liberan CO2. Estas funciones
metabólicas son empleadas para alimentar los sistemas de transporte activo que
mantiene la homeostasia iónica entre la célula y su medio (el glóbulo rojo y el
plasma). Su número varía entre los 4.5 a 6 millones por mm3 en el
varón y entre los 4 a 5.5 millones en la mujer, pero este número varía también
con la edad.
Los eritrocitos maduros carecen de núcleo pero tienen metabolismo,
consumen O2, ATP y glucosa y liberan CO2. Estas funciones
metabólicas son empleadas para alimentar los sistemas de transporte activo que
mantiene la homeostasia iónica entre la célula y su medio (el glóbulo rojo y el
plasma). Su número varía entre los 4.5 a 6 millones por mm3 en el
varón y entre los 4 a 5.5 millones en la mujer, pero este número varía también
con la edad.
Hematopoyesis:
Se
trata del proceso de formación de células sanguíneas. Todos los elementos
formes sanguíneos tienen un precursor común indiferenciado, llamado célula
madre hematopoyética pruripotencial (CMHP). A partir de esta célula madre se
forman dos precursores indiferenciados o multipotenciales que son la unidad
formadora de colonias del bazo (UFC-S) que es la célula madre de la línea
mieloide y la otra es la célula madre linfoide (CML), la célula madre de la
línea linfoidea.
Las
células madre se dividen continuamente pero lentamente y pueden
auto-regenerarse, lo que va a mantener una reserva de células madre en las
células óseas durante toda la vida del individuo, si bien disminuye con la
edad. Todos los elementos formes de la sangre excepto linfocitos se originan en
la médula ósea. Los linfocitos se originan en médula ósea pero también se
producen en respuesta a estímulos antigénicos en los tejidos linfoideos
periféricos.
En
la médula ósea roja las células madre multipotenciales se multiplican y se
diferencian en células comprometidas, en células prediferenciadas en una o dos
líneas celulares específicas. Las células prediferenciadas se dividen con
rapidez originando distintas líneas celulares. En nuestra médula ósea roja
existen cinco tipos de células prediferenciadas de la línea mieloide:
1. La unidad formadora de colonias
de eritrocitos.
2. La unidad formadora de colonias
de megacariocitos.
3. La unidad formadora de colonias
de eosinófilos.
4. La unidad formadora de colonias
de granulocitos y monocitos.
5. La unidad formadora de colonias
de basófilos.
Al microscopio no se diferencian
unas de otras pero si podemos si las dejamos crecer.
Lugares de formación de células sanguíneas:
Se
forman en diferentes lugares según sea la etapa embrionaria (hasta los tres
meses), la etapa fetal y la etapa después del nacimiento. En el primer mes se
forman en el saco vitelino. A partir de la quinta semana hasta el sexto mes
fundamentalmente en el hígado, pero también en el bazo y el timo. A partir del
sexto mes se forman en la médula ósea roja. Después del nacimiento todas las
células sanguíneas excepto los linfocitos se forman en la médula ósea roja.
Ahora
bien, en la niñez se producen en la médula ósea roja del esqueleto axial y en la
médula de los huesos de las extremidades, pero en la edad adulta se producen en
la médula del esqueleto axial y en los extremos proximales de fémur y húmero.
Factores necesarios para la
producción de células sanguíneas (hematopoyesis):
1. El factor de crecimiento.
2. El factor de diferenciación
(hormonas y citoquinas).
3. Es necesaria la eritropoyetina
para los glóbulos rojos y la trombopoyetina para las plaquetas.
Las
citoquinas son glicoproteínas producidas por diferentes tipos de células como
leucocitos macrófagos y células de la médula ósea. Hay dos familias de
citoquinas: por un lado los factores estimulantes de colonias (CFS) y las
interleuquinas que son muy numerosas (IL). Para la hematopoyesis se necesitan
también factores exógenos como el hierro, proteínas, lípidos y vitaminas.
Dentro de las vitaminas las más importantes son el ácido fólico, la B12
y B6.
Eritropoyesis:
No
se trata ni más ni menos que coger la célula madre y seguir su proceso. En la
médula ósea roja a partir de la célula madre multipotencial se forman unos
precursores prediferenciados, uno de ellos es el de eritrocitos.
Este
precursor comienza a diferenciarse dando lugar al proeritoblasto, que es la
primera célula de la línea eritropoyética reconocible. Los proeritoblastos se
dividen y diferencian y dan lugar al eritoblasto I, que es un eritoblasto
basófilo y donde comienza la síntesis de hemoglobia. El eritrocito I se vuelve
a dividir y diferenciar y se transforma en eritroblasto II que aún es basófilo
y donde continúa la síntesis de hemoglobina y aún presenta un núcleo definido.
Por lo tanto el eritoblasto II se divide y diferencia y da lugar al
policromatófilo. El policromatófilo ya contiene una gran cantidad de
hemoglobina, núcleo reducido y no se divide, solo se diferencia dando lugar al
acidófilo o normoblasto (eritoblasto ortocromático). Por lo tanto tiene mucha
hemoglobina con núcleo reducido poco definido, después se vuelve a diferenciar
y se transforma en un reticulocito que es considerado un glóbulo rojo inmaduro
porque ya no tiene núcleo, solo unos filamentos de cromatina y su hemoglobina
es la de un glóbulo rojo. El reticulocito sale a la sangre y a nivel óseo ya
pierde los filamentos de cromatina y se transforma en un glóbulo rojo maduro
que es que circula por la sangre.

En
condiciones normales (no patológicas) puede haber una pequeña cantidad de
reticulocitos circulantes pero para que sea normal el número de reticulocitos
tiene que estar entre el 0.5 y el 1 %
del total de glóbulos rojos. El glóbulo rojo maduro circula entre 90 y 120 días
realizando sus funciones. Cuando
finaliza este periodo el glóbulo rojo está envejecido y es eliminado,
aunque a veces la eritropoyesis puede ser ineficaz en alguna línea.
Cuantificación:
Recuento
de glóbulos rojos, el número de hematíes que vendrá dado siempre por un número
por 106/mm3. En segundo lugar debemos ver la cantidad de
hemoglobina en gramos por cada 100 ml de sangre, entre 14 y 16 gramos por 100
ml de sangre. En tercer lugar tenemos que ver el hematocrito en tanto %. A
partir de esto básico podemos calcular el volumen corpuscular medio, la
concentración corpuscular media de la hemoglobina que será la concentración de
hemoglobina en tanto % y podemos calcular la VSG (velocidad de sedimentación).
Funciones de los glóbulos rojos:
1. Transporte de O2 y CO2.
2. Aportan viscosidad de la sangre,
3. Actúa como amortiguador del pH,
fundamentalmente por la hemoglobina de la sangre.
Hemoglobina:
La
cantidad que representa es de 34 gramos por cada 100 ml de glóbulos rojos o lo
que es lo mismo, 16 g/100 ml de sangre en el hombre o 14 g/100 ml de sangre en
la mujer. La hemoglobina es el pigmento respiratorio de los vertebrados y está
formado por cuatro núcleos (pirroles) y un átomo de hierro. Además la
hemoglobina presenta cuatro cadenas polipeptídicas que en su conjunto llamamos
globina.

1. El ácido acético en el ciclo de
Krebs se transforma en succinil-CoA.
2. A continuación dos succinil-CoA
se unen a dos glicinas formando un pirrol.
3. Se unen cuatro pirroles y forman
un compuesto que es una protoporfirina, la protoporfirina IX.
4. Esta protoporfirina IX se une
con un hierro (Fe) y forma un grupo hemo.
5. Un grupo hemo se une a una
cadena polipeptídica del tipo α o β,
formando una cadena de hemoglobina α o β.
6. Se unen dos cadenas de hemoglobina α a dos cadenas
de hemoglobina β y tenemos la hemoglobina A.
La
globina está formada por cuatro cadenas polipeptídicas. La globina más común en
un adulto es la que está formada por dos cadenas α y dos
cadenas β. Las cadenas α tienen 141
aminoácidos y las cadenas β están compuestas por 149 aminoácidos. La hemoglobina formada por la
combinación de dos cadenas α y dos cadenas β es la
hemoglobina A, la más abundante en el adulto.
Pero esta
hemoglobina puede presentar alteraciones simplemente por el cambio de algún
aminoácido en sus cadenas. En este caso los eritroblastos que lleven
hemoglobinas anormales se desechan. En el feto la más abundante es otra
hemoglobina, la llamada hemoglobina fetal que no tiene cadenas β sino otro tipo
de cadenas denominadas cadenas γ. Esta hemoglobina fetal tiene una mayor
capacidad para captar O2, de tal modo que es ventajosa ya que puede
actuar a presiones parciales de O2 más reducida.
Metabolismo del hierro (Fe):
El hierro es
esencial para el transporte de O2 porque el O2 se une al
Fe de la hemoglobina. El Fe se absorbe en la primera parte del intestino
delgado por transporte activo y su absorción es más fácil en estado ferroso (Fe++)
que férrico (Fe+++, forma oxidada) y también se absorbe mejor en
estado hemínico (formando parte de grupos hemo) que no hemínicos.
En el
transporte activo participa una proteína transportadora, la ferroportina. El
transporte activo se produce en dos etapas: en primer lugar de la luz
intestinal a la mucosa intestinal y en la segunda etapa desde el interior de
esas células hasta el plasma. Este Fe absorbido o el Fe liberado por la
destrucción de los glóbulos rojos en el plasma nunca está libre sino que se
combina con una β-globulina que es la apotransferrina, formando un
un compuesto con el Fe llamado transferrina. Esta transferrina lleva al Fe
hacia la médula ósea (los eritroblastos) y lleva el Fe en exceso a los lugares
de depósito.
El Fe puede
depositarse en todas las células pero especialmente en las hepáticas, de hecho
es el lugar donde se almacenan el 60% del Fe en exceso. Al llegar al hígado el
Fe se libera de la apotransferrina y se combina con otra proteína que es la
apoferritina, formando un compuesto que es la ferritina, que es el Fe en
depósito. Esta ferritina cede la cantidad de Fe necesaria para su uso. El Fe
puede también almacenarse de una forma mucho más insoluble, llamada
hemosidenina, que forma grandes acúmulos en las células. Este tipo de
almacenamiento solo ocurre cuando la cantidad de Fe en el organismo es superior
al que puede contener la apoferritina.
La
transferrina tiene la capacidad de transportar el Fe al eritoblasto. En los
eritoblastos la transferrina se fija a los receptores de membrana y el
eritoblasto la ingiere por endocitosis junto con el Fe que lleva. De esta manera
llega hasta las mitocondrias del eritoblasto y a nivel mitocondrial descarga el
Fe que va a ser utilizado para la síntesis de grupos hemo. Por lo tanto queda
libre la apotransferrina que sale de nuevo al plasma por exocitosis.
Destrucción de glóbulos rojos:
Los glóbulos
rojos no sintetizan proteínas porque no presentan núcleo ni retículo
endoplasmático. Por lo tanto no son capaces de renovar sus enzimas ni los
componentes de membrana. Con el tiempo (90/120 días) se vuelven rígidos y
frágiles modificando sus proteínas de membrana. Al modificarse sus proteínas de
membrana se fijan inmunoglobulinas plasmáticas a esta membrana alterada y de
este modo son reconocidos y destruidos por los macrófagos. Fundamentalmente
estos macrófagos provienen del hígado, bazo, ganglios linfáticos y médula ósea,
pero la destrucción fisiológica del glóbulo roja puede ser también por
hemolisis extravascular (80-90%) o intravascular (10-20%).
La hemólisis
intravascular se produce porque al envejecer los glóbulos rojos va a entrar
agua en su interior, el eritrocito se hincha, la membrana se deteriora y deja
salir hemoglobina y otros constituyentes. Pero esta hemoglobina una vez que
pasa al plasma se va a unir a una proteína plasmática llamada aptoglobina y de
este modo va a ser transportada hasta que es captada por un macrófago.
La hemólisis
extravascular tiene lugar dentro de los macrófagos, bien porque llega la
hemoglobina de la intravascular o porque capta todo el glóbulo rojo. Dentro del
macrófago la hemoglobina se separa, lo primero en separarse es la globina, la
parte proteica, y quedan los grupos hemo. Esta globina se va a desdoblar en
aminoácidos que pasan a la circulación y son reutilizados. Los grupos hemo en
el interior del macrófago se rompen y dejan libre el Fe formando un compuesto,
un pigmento de color verde llamado biliverdina. Biliverdina que en el interior
del macrófago se reduce y forma un compuesto que es la bilirrubina.
Esta
bilirrubina pasa al plasma en donde se va a fijar con una albúmina que la
transporta a los hepatocitos del hígado. En el hígado se conjuga
fundamentalmente con glucurónico formando un compuesto que es la bilirrubina
conjugada (glucurónico de bilirrubina). Una pequeña parte de esta bilirrubina
conjugada puede pasar al plasma y va a ser eliminada por la orina, pero la
mayor parte de la bilirrubina conjugada
va a ser excretada a través del jugo biliar al intestinos. Pero en el
intestino a nivel de íleon y el colon la bilirrubina conjugada se enfrenta a la
flora bacteriana normal y por acción bacteriana se convierte en urobilinógeno.
Parte del urobilinógeno es absorbido a través de la pared intestinal y se va a
filtrar a nivel renal y será eliminado por la orina pero cuando se pone en
contacto con el oxígeno se oxida y se convierte en urobilina. La parte que
queda en el intestino se elimina a través de las heces.
Al
urobilinógeno en el intestino se le llama estercobilinógeno (a nivel del
colon). Este estercobilinógeno en contacto con el aire, con el oxígeno se oxida
y se transforma en estercobilina que es el responsable del color marrón de
nuestras heces. Parte del urobilinógeno se puede absorber por sangre portal
(vena porta) y llegar al hígado donde se transformaría en bilirrubina y pasaría
de nuevo al intestino a través de la bilis. Por lo tanto hay una recirculación
del urobilinógeno que pasa a nivel intestinal.
El glóbulo
rojo debido a que tiene hemoglobina y transporta CO2 y O2.
Un gramos de hemoglobina es capaz de transportar 1.33 cm3 de O2.
El glóbulo rojo posee un enzima llamado anhidrasa carbónica, que cataliza una
reacción:
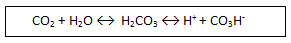
Es el mejor
tampón para la regulación del pH. Esto va a permitir que nuestra sangre
funcione en un momento determinado con una gran cantidad de CO2.
Sabeis más o menos cual sería la relación entre metabolismo del hierro y destrucción del glóbulo?
ResponderEliminargracias
Supongo que lo preguntarás porque es una de las posibles preguntas de medio interno. En cuanto a esta profesora se refiere, personalmente creo que lo mejor es, si ten manda relacionar dos apartados (como en este caso) escribirle todo lo que sepas de cada uno. En todo caso, la relación que yo creo que existe es que en la destrucción de los glóbulos rojos, al formar la biliverdina se desprende hierro (Fe) que podrá ser utilizado posteriormente para su metabolismo. Espero que te haya sido de ayuda :D
EliminarSí, justo lo que pensaba yo ;) Gracias!!
Eliminar